J. Mark Davis, Ph.D.
SPORTS SCIENCE EXCHANGE
GLUCIDES, ACIDES AMINÉS RAMIFIÉS ET ENDURANCE : L'HYPOTHÈSE DE LA FATIGUE CENTRALE
SSE n° 61, volume 9 (1996), numéro 2
Le présent article s'inspire d'un article paru dans un supplément de l'International Journal of Sports Nutrition, vol. 5, 1995. Ce supplément contient le compte-rendu de la conférence du Gatorade Sports Science Institute sur les aides ergogéniques, tenue à Chicago en novembre 1994.
J. Mark Davis
Department of Exercise Science
The University of South Carolina
Columbia, SC 29208
POINTS PRINCIPAUX
1. Les mécanismes de la fatigue centrale demeurent en grande partie inexplorés, mais d’après l’hypothèse de la fatigue centrale, une hausse de la sérotonine (5-HT) dans le cerveau peut entraîner une diminution de performance physique et sportive. Il y a maintenant suffisamment de preuves convaincantes pour affirmer qu’une augmentation du rapport tryptophane (TRP) plasmatique libre/acides aminés ramifiés (AAR), soit le rapport TRP/AAR, est associée à une hausse de la 5-HT dans le cerveau et à l’apparition de la fatigue pendant un exercice prolongé.
2. Avec l'administration de médicaments modifiant le taux de 5-HT du cerveau, les effets escomptés sur la performance physique sont effectivement observés. L'effet des manipulations nutritionnelles du rapport TRB/AAR sur la performance est moins bien connu.
3. Les effets des suppléments d’acides aminés ramifiés sur la performance physique sont mixtes et la méthodologie des études publiées laisse souvent à désirer. En revanche, lors d'un exercice prolongé, un apport en glucides entraîne de très importantes réductions du rapport TRP/AAR tout en améliorant la performance. Il est cependant difficile de distinguer les effets d'un apport en glucides sur les mécanismes qui se produisent dans le cerveau ou sur ceux qui se produisent dans les muscles eux-mêmes.
INTRODUCTION
La fatigue musculaire se définit communément comme l'incapacité de maintenir une force ou une puissance requise ou espérée (22). La fatigue musculaire est attribuable à certains déficits musculaires, notamment à la transmission de l’influx nerveux du muscle à la plaque motrice, puis à la propagation de cet influx à l’ensemble du muscle (35, 38), à l'interruption de la libération et de l'absorption de calcium dans le réticulum sarcoplasmique (23), à la déplétion du substrat (19) et à divers autres phénomènes métaboliques qui altèrent la production d’énergie et la contraction musculaire (27). La fatigue peut également résulter d’altérations internes du système nerveux central (SNC), même si les mécanismes sous-jacents inhérents à ce type de fatigue (fatigue centrale) ne sont presque pas connus.
Le rôle éventuel de la fatigue centrale pendant un exercice prolongé a suscité très peu d’intérêt de la part des scientifiques, même s’il est bien connu que des « facteurs psychologiques » peuvent modifier la performance physique (3). En fait, la fatigue ressentie par la plupart des gens au cours de leurs activités normales s’explique fort probablement par un manque d'activation des muscles sollicités par le SNC. Par ailleurs, une dysfonction des muscles eux-mêmes ne peut certainement pas expliquer la fatigue, souvent débilitante, qui accompagne les infections virales ou bactériennes, la récupération après une blessure ou une intervention chirurgicale, le syndrome de la fatigue chronique et la dépression. Dans ces cas précis, le SNC a probablement quelque chose à voir avec la fatigue, mais le phénomène n'a pas encore été élucidé.
De nombreuses études (3, 9, 34) soulignent le rôle important que joue la fatigue centrale pendant l’exercice chez les personnes en santé, mais la plupart de ces recherches n’ont pas réussi à proposer un mécanisme physiologique plausible. Il demeure que d’intéressantes nouvelles théories ont récemment été proposées et, selon ces théories, divers neurotransmetteurs comme la sérotonine (5-hydroxytryptamine ou 5-HT) (32), la noradrénaline et la dopamine (8, 28, 30, 33) participeraient à la fatigue centrale pendant un exercice. Le présent article porte avant tout sur les preuves scientifiques relatives à la 5-HT comme médiateur potentiel de la fatigue centrale pendant un exercice prolongé. Il sera également question d'intéressantes données scientifiques qui, quoique limitées, montrent qu'une alimentation appropriée pourrait modifier la synthèse de la 5-HT dans le cerveau et retarder la fatigue centrale. Aux fins du présent article, nous utiliserons une définition ad hoc de la fatigue centrale. Selon cette définition, la fatigue centrale constitue un sous-ensemble de la fatigue associée à des altérations particulières de la fonction cérébrale qui ne saurait s'expliquer logiquement par des marqueurs périphériques de la fatigue musculaire.
LA 5-HT DANS LE CERVEAU ET L’HYPOTHÈSE DE LA FATIGUE CENTRALE
En 1987, Newsholme et coll. (32) ont d’abord suggéré que la sérotonine pourrait agir comme médiateur de la fatigue centrale. De nombreuses publications associent les altérations de l’activité de la 5-HT à diverses réponses psychologiques comme l’éveil, la léthargie, la somnolence et l’humeur (42), qui pourraient toutes jouer un rôle dans les mécanismes centraux de la fatigue. De ce fait, la 5-HT est particulièrement intéressante comme facteur probable, car les mécanismes qui en contrôlent la synthèse et le métabolisme dans le cerveau sont susceptibles d’être modifiés par un exercice prolongé.
De façon générale, d’après l’hypothèse de la fatigue centrale, une hausse de 5-HT dans le cerveau peut entraver la fonction du SNC pendant un exercice prolongé et ainsi provoquer une diminution de performance physique et sportive (32). Il se produit une augmentation de la synthèse de la 5-HT dans le cerveau après une plus grande libération de tryptophane (TRP) provenant du sang, un acide aminé précurseur de la 5-HT. La plus grande partie du tryptophane plasmatique circulant est faiblement liée à l’albumine, mais c’est le tryptophane libre ou non lié qui traverse la barrière hémato-encéphalique. Ce transport du tryptophane se fait par un mécanisme particulier que d’autres grands acides aminés neutres utilisent, plus particulièrement les acides aminés ramifiés comme la leucine, l’isoleucine et la valine. De cette façon, la synthèse de la 5-HT augmente à mesure qu’augmente le rapport de la concentration de tryptophane (TRP) plasmatique libre à la concentration totale d’acides aminés ramifiés (AAR) dans le sang, ce qui se passe quand le rapport TRP/AAR augmente (15, 17, 18, 32). Il a été suggéré que cet effet se produirait pendant un exercice prolongé au moment où a) les acides aminés ramifiés sont absorbés par le sang et oxydés pour fournir de l’énergie aux muscles squelettiques et b) la concentration des acides gras libres (AGL) dans le sang augmente, ce qui entraîne une hausse simultanée du tryptophane plasmatique libre attribuable au fait que les AAR délogent le TRP de ses sites de liaison habituels sur les molécules d’albumine plasmatique (Figure 1).
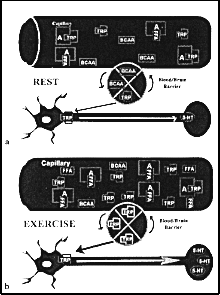
| Figure 1 : Principales composantes de l’hypothèse de la fatigue centrale, au repos et pendant un exercice prolongé. a) au repos, concentrations plasmatiques des acides aminés ramifiés (AAR), d’acides gras libres (AGL) et de tryptophane (TRP) libre et non lié à l’albumine et leurs effets possibles sur le passage du TRP à travers la barrière hémato-encéphalique pour que s'effectue la synthèse de la sérotonine (5-HT) dans les neurones sérotoninergiques. b) pendant un exercice prolongé, mécanisme permettant d’augmenter la synthèse de 5-HT dans le cerveau, entraînant ainsi la fatigue centrale. Adapté de J. D. Fernstrom. Dietary amino acids and brain function. J. Am. Diet. Dietary Assoc. 94:71-77, 1994. |
Les chercheurs ont d'abord vérifié la validité de cette hypothèse en faisant des essais chez l’humain et chez l’animal. Les premières études ont tenté de répondre à quelques questions fondamentales, dont les suivantes : Pendant un exercice prolongé, la fatigue est-elle associée à des hausses du taux de 5-HT et de son principal métabolite, l’acide 5-hydroxy-indole-acétique (5-HIAA) dans le cerveau, et ces hausses sont-elles attribuables à des augmentations du rapport TRP/AAR dans le sang ? Les altérations expérimentales de l’activité de la 5-HT dans le cerveau modifient-elles la fatigue associée à l’exercice sans effet apparent sur les marqueurs périphériques de la fatigue musculaire ? Les suppléments alimentaires peuvent-ils réduire l’augmentation du rapport TRP/AAR et ainsi améliorer l’endurance physique ? Il est à présent bien établi, même si les résultats sont en grande partie préliminaires, que les réponses aux deux premières questions viennent soutenir l’hypothèse de la fatigue centrale. Les résultats qui prouvent l’effet des stratégies nutritionnelles visant à réduire le taux de 5-HT dans le cerveau et ainsi retarder l’apparition de la fatigue sont moins convaincants.
ASSOCIATION ENTRE LA FATIGUE ET LES HAUSSES DE 5-HT ET DE 5-HIAA DANS LE CERVEAU
Les premières études chez l’animal ont été effectuées dans les laboratoires de recherche des professeurs Chaouloff (14-18) et Newsholme (13). Chaouloff et coll. (14, 15) ont fait courir des rats pendant 1 à 2 heures sur un tapis roulant (20 m/min). Cet exercice n'a eu aucun effet sur le tryptophane plasmatique total, mais les chercheurs ont observé une importante augmentation de tryptophane plasmatique libre, accompagnée d’une hausse de tryptophane dans le cerveau et d’une faible hausse, quoique significative, de 5-HIAA (le principal métabolite de la 5-HT). Ils ont par la suite montré que des hausses semblables de tryptophane et de 5-HIAA se produisent dans le liquide céphalorachidien (LCR) pendant l’exercice, et que les taux obtenus reviennent à leurs valeurs initiales environ une heure plus tard (17). Ces résultats, et d’autres obtenus par la suite, révèlent des augmentations de 5-HT et de 5-HIAA dans plusieurs régions du cerveau après une course sur tapis roulant de 90 minutes (16); ce sont les premiers résultats confirmant l’hypothèse selon laquelle une course d’endurance chez le rat peut augmenter la synthèse et le renouvellement de la 5-HT dans le cerveau. Ils ont également montré, de façon plutôt élégante, que l’augmentation de tryptophane plasmatique libre est le principal responsable de cette réponse. Toutefois, comprendre comment ces résultats expliquent les mécanismes de la fatigue n'a pas fait l'objet de leur recherche.
Blomstrand et coll. (13), dans leur étude chez des rats courant jusqu’à épuisement sur un tapis roulant, montrent une augmentation du tryptophane plasmatique libre (mais non du tryptophane total) et des concentrations de tryptophane, de 5-HT et de 5-HIAA dans certaines régions du cerveau, tant chez les rats entraînés (course d’environ 180 min) que chez les rats non entraînés (course d’environ 72 min). Nous avons élargi ces résultats en procédant à une étude sur les effets des changements du taux de 5-HT et de dopamine dans le cerveau sur le temps de course, la dopamine étant un neurotransmetteur ayant un rôle important dans la motivation, l’état de veille et le contrôle neuromusculaire pendant une course d’endurance jusqu’à épuisement (6). Les taux de 5-HT et de dopamine ainsi que de leurs principaux métabolites, le 5-HIAA et l’acide 3,4-dihydroxyphénylacétique (DOPAC) dans le mésencéphale, le striatum, l’hypothalamus et l’hippocampe de rats sacrifiés ont été mesurés au repos, après une heure d’effort sur tapis roulant et à l’état d’épuisement, soit après environ 3 heures. La vitesse et l’inclinaison du tapis roulant étaient réglées à environ 60 à 65 % du VO2max des rats (20 m min-l, % d'inclinaison). Après une heure d'exercice, les taux de 5-HT et de 5-HIAA avaient augmenté dans toutes les régions du cerveau, sauf dans l’hippocampe, où seulement le taux de 5-HIAA était plus élevé. Le taux de 5-HT dans le cerveau est demeuré élevé à l’épuisement, tandis que le 5-HIAA avait encore davantage augmenté dans le mésencéphale et le striatum. Les taux de dopamine et de DOPAC ont également augmenté dans le mésencéphale, le striatum et l’hypothalamus après une heure, mais sont retombés aux valeurs initiales à l’épuisement. Ces résultats indiquent que l’activité de la 5-HT augmente dans le cerveau pendant un exercice prolongé et semble atteindre son maximum à l’épuisement. Fait à noter, l’activité de la dopamine dans le cerveau, activité généralement associée à une augmentation de la vigilance et de la coordination musculaire (24), a effectivement diminué vers la fin de l’exercice prolongé, soit au fur et à mesure que la fatigue se faisait sentir. La signification de cette apparente relation inverse entre d’éventuels effets suppresseurs de la 5-HT dans le cerveau et les effets stimulants de la dopamine est intéressante en ce qui concerne la fatigue centrale, mais mérite d’être approfondie.
ALTÉRATIONS EXPÉRIMENTALES DE L'ACTIVITÉ DE LA 5-HT DANS LE CERVEAU
Même s’il semble y avoir une bonne relation entre un taux élevé de 5-HT dans le cerveau et la fatigue induite par l’exercice, il serait prématuré d'en conclure que les altérations de l’activité de la 5-HT dans le cerveau causent la fatigue centrale. Afin de mieux explorer cette question de cause à effet, nous avons effectué une série d’essais pour déterminer si certaines modifications dans l'activité de la 5-HT dans le cerveau après l'administration de certains médicaments pouvaient influer sur l'endurance physique des rats. Selon notre hypothèse, si une augmentation de 5-HT dans le cerveau est responsable de la fatigue centrale lors d'un exercice prolongé, l’administration de médicaments connus pour augmenter l’activité de la 5-HT (agonistes de la 5-HT) devrait provoquer rapidement une fatigue, tandis que des médicaments connus pour diminuer l’activité de la 5-HT (antagonistes de la 5-HT) devraient retarder la fatigue. Dans un essai préliminaire, après l'administration de différentes doses d’un agoniste des récepteurs de la 5-HT1c, le métachlorophénylpipérazine (mCPP), une diminution liée à la dose du temps d’effort jusqu’à épuisement a été observée (5). Dans l’essai suivant, un agoniste de la 5-HT plus générique, le dimaléate de quipazine, ou un antagoniste de la 5-HT, le LY-53857 ont été administrés (4). L’agoniste de la 5-HT a de nouveau provoqué une diminution liée à la dose du temps jusqu’à épuisement, tandis que l’antagoniste de la 5-HT a retardé la fatigue. L'hypothèse selon laquelle ces effets induits par des médicaments découlent de fonctions cérébrales altérées est corroborée par l'observation voulant que la fatigue ne puisse s’expliquer par un changement dans la température du corps, la glycémie, le glycogène musculaire ou hépatique, ni même des différentes hormones de stress (6).
Ces résultats, obtenus à l'aide d'une approche pharmacologique propre au rat, ont récemment été confirmés par deux études chez l’humain. Dans ces études, l’activité de la 5-HT dans le cerveau a été augmentée par l’administration de paroxétine (41) ou de fluoxétine (20), deux inhibiteurs du recaptage de la 5-HT à partir des terminaisons nerveuses, dont l’utilisation est approuvée chez l’humain et qui, lors d'une administration aiguë, agissent comme des agonistes de la 5-HT, étant donné qu’ils inhibent son retrait. Quand ces médicaments étaient administrés avant un effort prolongé lors d'une course à pied ou d'une épreuve de cyclisme à 70 % du VO2max, le temps jusqu’à épuisement était plus court (41) et l’intensité de l’effort perçu était plus élevée (20) qu’avec l’administration d’un placebo. Les sujets n’ont signalé aucun effet secondaire inhabituel, et aucune différence dans les divers marqueurs des fonctions cardiovasculaires, thermorégulatrices ou métaboliques n'a été observée entre médicament et placebo.
Les études citées ci-dessus chez le rat et l’humain semblent nettement démontrer que l’activité de la 5-HT dans le cerveau augmente pendant un exercice prolongé et qu'elle pourrait être la cause de la fatigue centrale. Toutefois, la validité de ces résultats continuera d'être remise en question jusqu’à ce que de nouvelles méthodes permettent de mesurer plus directement la fatigue centrale lors d’un exercice dynamique chez l’humain et jusqu’à ce que nous en sachions plus sur les mécanismes physiologiques à l’origine d’un tel effet.
Les chercheurs ne font que commencer à explorer les éventuels mécanismes physiologiques sous-jacents à l’effet éventuel d’une augmentation de l’activité de la 5-HT sur la fatigue centrale. Le système sérotoninergique est associé à de nombreuses fonctions cérébrales qui peuvent avoir une influence positive ou négative sur l’endurance physique. Une augmentation de l'activité sérotoninergique peut induire la fatigue en inhibant le système dopaminergique (6, 18) ou en réduisant la vigilance et la motivation nécessaires pour performer (29,42). De plus, l’activité sérotoninergique peut modifier l’activité de l’axe hypothalamo-hypophyso-surrénalien, la thermorégulation, la douleur et l’humeur en fonction d’une situation donnée et de l’espèce à l'étude (1, 2, 42). D’après nos observations, lors d'un exercice prolongé effectué par un rat, la fatigue est associée à une augmentation de l’activité de la 5-HT et à une réduction de la dopamine dans le cerveau (5, 6); notre hypothèse de travail est donc qu'un faible rapport 5-HT/dopamine dans le cerveau améliore la performance (c.-à-d. augmentation de la vigilance et de la motivation, et coordination neuromusculaire optimale), tandis qu’un rapport 5-HT/dopamine élevé diminue la performance (p. ex., diminution de la motivation, léthargie, fatigue et baisse de la coordination motrice), ce qui constitue une fatigue centrale.
EFFETS NUTRITIONNELS DE LA 5-HT SUR LA FATIGUE CENTRALE
Pour des raisons éthiques évidentes, c'est chez le rat que les chercheurs ont étudié les effets de la fatigue sur les concentrations de 5-HT et de ses métabolites dans certaines régions du cerveau. Les recherches chez l’humain ont surtout porté sur les facteurs nutritionnels qui modifient la disponibilité du tryptophane pour le cerveau, c’est-à-dire des marqueurs hypothétiques de la fatigue centrale.
Blomstrand et coll. (10) ont été les premiers à analyser la question chez l’humain. Dans un premier temps, ils ont analysé la concentration de tryptophane plasmatique libre chez 22 sujets avant et après un marathon et ont constaté qu'elle était environ 2,4 fois plus élevée après la course tandis que les acides aminés ramifiés (AAR) étaient légèrement moins élevés (-19 %). Ils ont observé le même genre de réponse après une partie de soccer (augmentation de 45 % du TRP; diminution de 29 % des AAR) et après une épreuve de ski de fond prolongée (diminution de 28 % des AAR; valeur de TRP non relevée) (11, 12). Dans ces études, la baisse du rapport TRP/AAR après un exercice a confirmé leur hypothèse selon laquelle l’exercice prolongé augmente la disponibilité du TRP pour le cerveau et qu’il peut s’ensuivre une augmentation de l’activité de la 5-HT ainsi qu'une fatigue centrale. Ces résultats ont servi de fondement à leurs théories voulant que d’éventuelles stratégies nutritionnelles puissent favoriser un retard de la fatigue centrale pendant un exercice prolongé (11-13).
La possibilité qu'en théorie la fatigue centrale puisse être retardée par des stratégies nutritionnelles modifiant le rapport TRP/AAR est particulièrement intéressante. Les recherches ont porté sur deux principales stratégies faisant appel à des suppléments d’acides aminés ramifiés, de glucides, ou les deux, lors d'un exercice. D’un point de vue théorique, ces deux stratégies permettraient de diminuer le rapport TRP/AAR et ainsi réduire la disponibilité du TRP pour la synthèse de la 5-HT dans le cerveau.
Blomstrand, Newsholme et coll. ont privilégié l’administration d’acides aminés ramifiés comme moyen pour retarder la fatigue centrale. Ils rapportent que l’administration de 7,5 à 21 g d’acides aminés ramifiés avant et pendant un marathon, une compétition de ski de fond ou une partie de soccer est associée à de faibles améliorations de la performance physique (11) ou mentale (12) chez certains sujets. Toutefois, si ces études sur le terrain cherchaient à reproduire les situations réelles que vivent les athlètes, elles ont souvent peu de valeur du point de vue scientifique. Par exemple, il arrive souvent que la répartition des les sujets ne soient pas bien faite entre le groupe témoin et le groupe expérimental avant l'exercice; une telle répartition est pourtant utile pour éviter de trop grandes différences associées au conditionnement physique, à l'entraînement, à la composition corporelle ou à d'autres facteurs propres à chaque sujet, d'un groupe à l'autre. De plus, il arrive souvent que les études de cette nature ne soient pas faites, ou ne peuvent être faites, « à l’insu » des participants pour ce qui est des traitements expérimentaux afin d'éviter que certains sujets aient des préjugés en faveur d'un traitement plutôt qu'un autre. Enfin, ces études omettent souvent de contrôler d’importantes variables comme l’intensité de l’effort ainsi que l’apport en aliments et en eau dans chaque groupe de traitement. Ces limites et bien d’autres augmentent la probabilité que les effets bénéfiques attribués à un supplément alimentaire en particulier découlent en fait d’idées préconçues de la part des sujets, de différences inhérentes à chacun des groupes à l'étude et d’une ou de plusieurs variables non contrôlées.
Le scepticisme suscité par les résultats de ces premières études sur le terrain s'est intensifié après la divulgation des résultats obtenus dans de récentes études bien contrôlées effectuées en laboratoire et selon lesquelles les suppléments d’acides aminés ramifiés n’ont aucun effet positif sur la performance lors d'un exercice d'endurance. Varnier et coll. (36) ont effectué une perfusion d’environ 20 g d’acides aminés ramifiés ou de solution saline pendant 70 minutes avant l’exercice lors d'une étude croisée à double insu. Ils n'ont observé aucune différence de performance lors d'un test d’effort progressif jusqu’à épuisement. Verger et coll. (37) rapportent également que l’administration à des rats de grandes quantités d’acides aminés ramifiés (comparativement à de l’eau ou à du glucose) a, en réalité, provoqué une fatigue précoce lors d’une épreuve de course prolongée sur tapis roulant.
Il est important de souligner que l'administration de grandes quantités d’acides aminés ramifiés nécessaires pour obtenir des changements pertinents sur le plan physiologique dans le rapport TRP/AAR risque d'augmenter les taux d’ammoniaque, qui peuvent s’avérer toxiques pour le cerveau et avoir des effets négatifs sur le métabolisme musculaire (7, 39, 40). La toxicité aiguë de l’ammoniaque, bien que transitoire et réversible, peut se révéler suffisamment grave dans les régions cruciales du système nerveux central pour altérer la performance (coordination, contrôle moteur) ou provoquer de graves symptômes de fatigue centrale, ou les deux (7). L'accumulation d'ammoniaque pourrait également provoquer la fatigue des muscles sollicités en épuisant les squelettes de carbone (pyruvate) issus de la glycolyse et en drainant les intermédiaires du cycle des acides tricarboxyliques, qui sont liés à la production de glutamine par des réactions de transamination (39, 40), ce qui pourrait vraisemblablement nuire au métabolisme de l’oxydation dans les muscles et entraîner une fatigue précoce.
Pour cette raison et parce que l'administration de fortes doses d’acides aminés ramifiés pendant l’exercice risque de ralentir l’absorption d’eau dans les intestins et de provoquer des troubles gastro-intestinaux, nous avons effectué une étude croisée à double insu afin de déterminer l’effet d’une dose plus faible d’acides aminés ramifiés (environ 0,5 g h-1 sous forme de boisson contenant des glucides et des électrolytes), au goût plus agréable, lors d'une épreuve de cyclisme à 70 % du VO2max et jusqu’à épuisement (25). Cette faible dose d’acides aminés ramifiés (AAR) devait remplacer la quantité maximale d'AAR qui serait probablement absorbée et métabolisée par les muscles dans ces conditions et pour réduire le risque que les suppléments d’AAR réduisent les taux d’absorption d’eau dans les intestins et entraînent des malaises gastro-intestinaux ou, tout simplement, pour lui donner du goût. Les résultats de cette étude ont montré que de faibles quantités d’AAR ont bon goût quand ils sont ajoutés à une boisson contenant des glucides et des électrolytes, qu’ils ne causent pas de malaises gastro-intestinaux et qu’ils empêchent la légère baisse des taux d’AAR plasmatiques associée à une séance de cyclisme prolongée, ce qui n'avait pas été le cas quand les sujets avaient bu une boisson contenant des glucides et des électrolytes, mais sans suppléments d’AAR. Toutefois, les suppléments d’AAR ajoutés n’ont pas amélioré le temps jusqu'à épuisement, n’ont pas atténué l’intensité de l’effort perçu, ni modifié les divers paramètres des fonctions cardiovasculaires et métaboliques.
Nous en avons déduit qu'un apport en glucides serait probablement une meilleure stratégie pour retarder la fatigue étant donné qu'un tel apport entraînerait une forte diminution du taux de tryptophane plasmatique libre et du rapport TRP/AAR pendant l’exercice, sans les effets négatifs associés aux fortes doses d’AAR, et cela en raison des effets suppresseurs clairement établis d'un apport en glucides sur la mobilisation des acides gras libres, qui tentent de se lier aux mêmes sites que le tryptophane sur les molécules d’albumine plasmatique. Nous avons donc suggéré qu'un apport en glucides pourrait réduire les concentrations de tryptophane plasmatique libre et le rapport TRP/AAR ce qui, en retour, pourrait interrompre la production de 5-HT dans le cerveau, minimisant ainsi la fatigue centrale (21). Ces effets se produiraient en plus des effets bénéfiques bien connus des suppléments de glucides sur les mécanismes périphériques de la fatigue (19).
Pour vérifier cette hypothèse, nous avons effectué en laboratoire une étude à double insu, au cours de laquelle les sujets devaient boire 5 mL/kg-1 h-1 d'un placebo constitué d'eau, une boisson contenant 6 % de glucides et d'électrolytes ou une boisson contenant 12 % de glucides et d'électrolytes pendant une séance de cyclisme à 70 % du VO2 max jusqu’à épuisement (21). Chez les sujets ayant bu le placebo, le taux de tryptophane plasmatique libre a été multiplié environ par sept (de façon directement proportionnelle aux taux d’acides gras libres dans le sang), tandis que les taux de tryptophane total et d'acides aminés ramifiés ont été très peu modifiés pendant la séance de cyclisme. Chez les sujets ayant bu les boissons contenant 6 % ou 12 % de glucides et d'électrolytes, le taux de tryptophane plasmatique libre a très peu augmenté et la fatigue a été retardée d’environ une heure. Les apports en glucides ont entraîné une légère baisse des AAR dans le sang (environ 19 % et 31 %, respectivement, pour les groupes ayant bu la boisson à 6 % ou à 12 % de glucides et d'électrolytes), mais cette réduction est probablement négligeable en comparaison de la très forte diminution (de cinq à sept fois) du tryptophane plasmatique libre (20). Même s'il n'a pas été possible de faire la distinction entre les effets bénéfiques de l'apport en glucides en fonction des mécanismes centraux et périphériques de la fatigue, il est intéressant de noter que le retard marqué de la fatigue ne pouvait être expliqué par les marqueurs typiques de la fatigue musculaire périphérique à laquelle participent les fonctions cardiovasculaires, thermorégulatrices et métaboliques.
RÉSUMÉ ET CONCLUSIONS
Lors d'un exercice prolongé, la fatigue est traditionnellement associée aux mécanismes qui entraînent une dysfonction des processus de contraction musculaire. Plus récemment, toutefois, l’intérêt envers les mécanismes centraux de la fatigue a augmenté à mesure que les rouages physiologiques du système nerveux central étaient mieux compris. Malheureusement, les progrès dans ce domaine ont été freinés par le manque de bonnes pratiques méthodologiques permettant de faire, chez l'humain, la distinction entre mécanismes centraux et mécanismes périphériques de la fatigue lors d’un exercice dynamique mobilisant tout l’organisme.
De plus en plus de faits probants se dégagent toutefois quant au rôle de la 5-HT dans d'apparition de la fatigue centrale lors d'un exercice prolongé, même si ses mécanismes précis n’ont pas été établis. Les études révèlent que a) les concentrations de la 5-HT et de son principal métabolite, le 5-HIAA, augmentent dans plusieurs régions du cerveau pendant un exercice prolongé et atteignent leur valeur maximale à l’épuisement; b) l’augmentation de la synthèse et du renouvellement de la 5-HT dans le cerveau est presque certainement le résultat d’une augmentation du taux de tryptophane plasmatique libre et du rapport TRP/AAR; et c) l’administration de médicaments agonistes ou antagonistes de la 5-HT peut faire diminuer ou augmenter le temps jusqu'à épuisement en l’absence de tout marqueur périphérique de la fatigue musculaire.
Il y a de bonnes raisons de croire qu’une alimentation adéquate pourrait jouer un rôle dans le retard de la fatigue pendant un exercice prolongé, mais les données scientifiques en ce domaine sont beaucoup moins convaincantes. Peu d’études ont porté sur le rôle supposé des suppléments d’AAR, et il y a lieu de penser que cette approche pourrait ne pas être viable. En revanche, les suppléments de glucides sont liés à de fortes diminutions du tryptophane plasmatique libre et du rapport TRP/AAR, et la fatigue est, de toute évidence, retardée par cette stratégie nutritionnelle. Dans ce cas, cependant, il est impossible de distinguer avec certitude entre les effets d'un apport en glucides sur les mécanismes de la fatigue et les effets bénéfiques bien établis des suppléments de glucides sur la contraction musculaire.
La possibilité qu'il existe des liens entre nutrition, neurochimie du cerveau et performance sportive est emballante, et il pourrait bien en émerger un tout nouveau volet en matière de recherche sur la nutrition sportive. Toutefois, même si les preuves obtenues sont fascinantes et qu'elles reposent sur un raisonnement intuitif qui soit logique, nos connaissances en ce domaine sont, au mieux, rudimentaires.
References
1. Akil, H., and J.C. Liebeskind. Monoaminergic mechanisms of stimulation-produced analgesia. Brain Res. 94:279-296, 1975.
2. Alper, R.H. Evidence for central and peripheral serotonergic control of corticosterone secretion in the conscious rat. Neuroendocrinology 51:255-260, 1990.
3. Asmussen, E. Muscle fatigue. Med. Sci. Sports. 11(4): 313-321, 1979.
4. Bailey, S.P., J.M. Davis, and E.N. Ahlborn. Effect of increased brain serotonergic (5-HT,c) actiyity on endurance performance in the rat. Acta Physiol. Scand. 145 (91): 75-76, 1992.
5. Bailey, S.P., J.M., Davis, and E.N. Ahlborn. Brain serotonergic activity affects endurance performance in the rat. Int. J. Sports Med. 6:330-333, 1993.
6. Bailey, S.P., J.M. Davis, and E.N. Ahlborn. Neuroendocrine and substrate responses to altered brain 5-HT activity during prolonged exercise to fatigue. J. Appl. ?hysiol. 74(6):3006-3012, 1993.
7. Banister, W.W., and B.J.C. Cameron. Exercise-induced hyperammonemia: Peripheral and central effects. Int. J. Sports Med. 11 (Suppl. 2):S129-S142, 1990.
8. Bhagat, B., and N. Wheeler. Effect of amphetamine on the swimming endurance of rats. Neuropharmacology, 12:711-713, 1973.
9. Bigland-Ritchie, B., F. Furbush, and J.J. Woods. Fatigue of intermittent submaximal voluntary contractions: Central and peripheral factors. J. Appl. Physiol. 61 (2):421-429, 1986.
10. Blomstrand, E., F. Celsing, and E.A. Newsholme. Changes in plasma concentrations of aromatic and branch-chain amino acids during sustained exercise in man and their possible role in fatigue. Acta Physiol. Scand. 133:115 - 121, 1988.
11. Blomstrand, E., P. Hassmen, B. Ekblom, and E.A. Newsholme. Administration of branched-chain amino acids during sustained exercise--Effects on performance and on plasma concentration of some amino acids. Eur. J. Appl. Physiol. 63:83-88, 1991.
12. Blomstrand, E., P. Hassmen, and E.A. Newsholme. Effect of branched-chain amino acid supplementation on mental performance. Acta Physiol. Scand. 136:473-481, 1991.
13. Blomstrand, E., D. Perrett, M. Parry-Billings, and E.A. Newsholme. Effect of sustained exercise on plasma amino acid concentrations and on 5-hydroxytryptamine metabolism in six different brain regions in the rat. Acta Physiol. Scand. 136:473-481, 1989.
14. Chaouloff, F., J.L. Elghozi, Y. Guezennec, and D. Laude. Effects of conditioned running on plasma, liver and brain tryptophan and on brain 5-hydroxytryptamine metabolism of the rat. Brit. J. Pharm. 86:33-41, 1985.
15. Chaouloff, F., G.A. Kennett, B. Serrurier, D. Merina, and G. Curson. Amino acid analysis demonstrates that increased plasma free tryptophan causes the increase of brain tryptophan during exercise in the rat. J. Neurochem. 46:1647-1650, 1986.
16. Chaouloff, F., D. Laude, and J.L. Elghozi. Physical exercise: Evidence for differential consequences of tryptophan on 5-HT synthesis and metabolism in central serotonergic cell bodies and terminals. J. Neural Trans. 78:121-130, 1989.
17._ roxyindoleacetic acid, and homovanillic acid in ventricular cerebrospinal fluid of the conscious rat. J. Neurochern. 46:1313-1316, 1986.
18. Chaouloff, F., D. Laude, D. Merino, B. Sermrier, Y. Guezennec, and J.L. Elghozi. Amphetamine and alpha-methyl-p-tyrosine affect the exercise induced imbalance between the availability of tryptophan and synthesis of serotonin in the brain of the rat. Neuropharmacology 26:1099-l 106, 1987.
19. Coggan, A.R., and E.F. Coyle. Carbohydrate ingestion during prolonged exercise: Effects on metabolism and performance. Exer. Sport Sci. Rev. 19: 1-40, 1991.
20. Davis, J.M., S.P. Bailey, D.A. Jackson, A.B. Strasner, and SIL. Morehouse. Effects of a serotonin (5-HT) agonist during prolonged exercise to fatigue in humans. Med. Sci. Sports Exerc. 25(5):$78, 1993.
21. Davis, J.M., S.P. Bailey, J.A. Woods, F.J. Galiano, M. Hamilton, and W.P. Bartoll. Effects of carbohydrate feedings on plasma free-tryptophan and branched-chain amino acids during prolonged cycling. Eur. J. Appl. Physiol. 65:513-519, 1992.
22. Enoka, R.M., and D.G. Stuart. Neurobiology of muscle fatigue. J. Appl. Physiol. 72(5): 1631-1648, 1992.
23. Fitts, R.H., and J.M. Metzget. Mechanisms of muscular fatigue. In Principles of Exercise Biochemistry (2nd ed., Vol. 38), J.R. Poortmans (Ed.), Basel, Switzerland: Karger, 1993, pp. 248-268.
24. Freed, C.R., and B.K. Yamamoto. Regional brain dopamine metabolism: A marker for speed, direction, and posture of moving animals. Science 229:62-65, 1985.
25. Galiano, F.J., J.M. Davis, S.P. Bailey, J.A. Woods, and M. Hamilton. Physiologic, endocrine and performance effects of adding branch chain amino acids to a 6% carbohydrate-electrolyte beverage during prolonged cycling. Med. Sci. Sports Exerc. 23(4):S14, 1991.
26. Gerald, M.C. Effect of (+)- amphetamine on the treadmill endurance performance of rats. Neuropharmacology 17:703-704, 1978.
27. Green, HJ. Neuromuscular aspects of fatigue. Can. J. Sports Sci. 12 (Suppl. 1):7s-19s, 1987.
28. Heyes, M.P., E.S. Gamett, and G. Coates. Central dopaminergic activity influences rats' ability to exercise. Life Sci. 36:671-677, 1985.
29. Jauvet, M. and J-F. Pujol. Effects of central alterations of serotonergic neurons upon the sleep-waking cycle. Adv. Blochem. Psychopharm. 11:199-209, 1974.
30. Laties, V.G., and B. Weiss. The amphetamine margin in sports. Federation Proc. 40:2689-2692, 1981.
31. Leathwood, P.D., and J.D. Ferustrom. Effect of an oral tryptophardcarbohydrate load on tryptophan, large neutral amino acid, and serotonin and 5-hydroxyindoleacetic acid levels in monkey brain. J. Neural Tran. 79:25-34, 1990.
32. Newsholme, E.A., I.N. Acworth, and E. Blomstrand. Amino acids, brain neurotransmitters and a functional link between muscle and brain that is important in sustained exercise. In Advances in Mvochemistry, G. Benzi (Ed.), London: John Libbey Eurotext, 1987, pp. 127-133.
33. Salter, C.A. Dietary tyrosine as an aid to stress resistance among troops. Military Med. 154: 144-146, 1989.
34. Secher, N.H. Central nervous influence on fatigue. In Endurance in Sport, R.J. Shephard and P.-O. Astrand (Eds.), Boston: Blackwell Scientific, 1992, pp. 96-106.
35. Sjogaard, G. Muscle fatigue. Med Sports Sci. 26:98-109, 1987.
36. Varnier, M., P. Sarto, D. Martines, L. Lora, F. Carmignoto, G. Leese, and R. Naccarato. Effect of infusing branched-chain amino acid during incremental exercise with reduced muscle glycogen content. Eur. J. Appl. Physiol. 69:26-31, 1994.
37. Verger, P.H., P. Aymard, L. Cynobert, G. Anton, and R. Luigi. Effects of administration of branched-chain amino acids vs. glucose during acute exercise in the rat. Physiol. Behav. 55(3):523-526, 1994.
38. Vollestad, N.K., and O.M. Sejersted. Biochemical correlates of fatigue. Eur. J. Appl. Physiol. 57:336-347, 1988.
39. Wagenmakers, A.J.M., E.J. Bechers, F. Brouns, H. Kuipers, P.B. Soeters, G.J. Van der Vusse, and W.H.M. Saris. Carbohydrate supplementation, glycogen depletion, and amino acid metabolism during exercise. Am. J. Physiol. 260:E883-E890, 1991.
40. Wagenmakers, A.J.M., J.H. Coakley, and R.H.T. Edwards. Metabolism of branched-chain amino acids and ammonia during exercise: Clues from McArdle's disease. Int. J. Sports Med. 11:S101-S113, 1990.
41. Wilson, W.M., and R.J. Maughan. Evidence for a possible role of 5-hydroxytryptamine in the genesis of fatigue in man: Administration of paroxetine, a 5-HT re-uptake inhibitor, reduces the capacity to perform prolonged exercise. Exp. Physiol. 77:921-924, 1992.
42. Young, S.N. The clinical psychopharmacology of tryptophan. In Nutrition and the Brain (Vol. 7), R.J. Wurtman and J.J. Wurtman (Eds.), New York: Raven, 1986, pp. 49-88.