POINTS IMPORTANTS
L’exercice et l'alimentation influent sur le développement du cerveau chez l'enfant.
L’exercice et l'alimentation peuvent ralentir et inverser la perte cognitive chez les personnes âgées.
L’exercice et l'alimentation influent sur la santé du cerveau grâce à divers mécanismes au cours desquels naissent de nouveaux neurones (neurogenèse).
Pendant un exercice, la fatigue peut être induite par le cerveau.
Des interventions en nutrition peuvent influer sur l’incidence d'une « fatigue centrale ou cérébrale » pendant l'exercice.
INTRODUCTION
L’activité physique est associée à une réduction de nombreux troubles physiques et mentaux. D'innombrables études ont en effet montré que l’activité physique peut réduire l’incidence des maladies cardiovasculaires, des cancers du côlon et du sein, de l'obésité, ainsi que d'autres problèmes de santé comme la maladie d’Alzheimer, la dépression et l’anxiété (Gómez-Pinilla, 2011; Van Praag, 2009). Un grand nombre d'études prospectives et transversales montrent qu'une alimentation pouvant favoriser la fonction cognitive chez les personnes âgées devrait contenir plusieurs portions de poisson (2 à 5) par semaine et plusieurs portions par jour de céréales, de fruits aux couleurs vives ou foncées et des légumes verts (Parrott &Greenwood, 2007). C’est pourquoi l’alimentation et l’exercice font partie des interventions en nutrition visant à inverser les effets négatifs potentiels du vieillissement sur les fonctions cérébrales. Le présent article décrit de quelle façon l’exercice et l'alimentation peuvent influer sur le développement du cerveau, les fonctions mentales et cognitives (p. ex., se rappeler certaines choses, comme l’endroit où on a garé sa voiture à l’aéroport).
LA VALEUR DE L’EXERCICE ET DE L'ALIMENTATION POUR LE CERVEAU
Enfants - Un cerveau en développement
Il est de plus en plus reconnu que les enfants d'aujourd'hui grandissent en étant inactifs et en mauvaise condition physique, et que ces facteurs de leur mode de vie sont associés à l’apparition précoce de plusieurs maladies chroniques, dont le diabète de type 2 et l’obésité. De nombreuses études transversales et longitudinales ont révélé qu'il existe un lien entre l'embonpoint et de piètres résultats scolaires. La capacité aérobique est aussi liée de façon positive aux fonctions cognitives et à la réussite scolaire (Davis et Cooper, 2011). Plusieurs études semblent indiquer que les capacités cognitives et les résultats scolaires des enfants’ peuvent être affectés par leur condition physique générale (Davis et Cooper, 2011).
L’alimentation peut également influer à la fois sur le développement et sur la santé des structures et des fonctions du cerveau. Une saine alimentation fournit les éléments de base au cerveau pour créer et conserver les liaisons neuronales essentielles à l’amélioration des fonctions cognitives et des résultats scolaires. Les facteurs alimentaires ont de nombreux effets positifs sur les fonctions et la plasticité neuronales. Par exemple, les acides gras oméga-3 participent à la structure même du cerveau. Ils sont essentiels pour permettre la signalisation intercellulaire et, par conséquent, pour exercer une influence positive sur la fonction synaptique. Toutefois, les régimes riches en sucres, en lipides saturés ou très caloriques sont considérés comme néfastes pour les fonctions neuronales puisqu’ils réduisent la plasticité synaptique et les fonctions cognitives Gómez-Pinilla, 2011). La fonction cérébrale dépend évidemment d’une alimentation adéquate, et les variations à court terme dans la quantité et la composition de l'apport nutritionnel chez les personnes en santé influent sur leur fonction cognitive. Des études ont révélé que le petit-déjeuner est aurait de nombreux effets positifs sur la fonction cognitive des enfants bien nourris (Hoyland et coll., 2009; Taki et coll., 2010). Il a été montré que l’exercice interagit avec les modifications alimentaires —augmentant ainsi ses effets positifs sur la fonction cérébrale et diminuant en même temps les effets nocifs d’un régime alimentaire trop riche en matières grasses. Dans l'ensemble, les preuves semblent indiquer que des stratégies axées à la fois sur l’exercice et un régime alimentaire peuvent favoriser de façon maximale la santé des nerfs. De plus, Pivik et coll. (2012) ont récemment analysé l’effet du repas matinal sur les fonctions mentales complexes des enfants (de 8 à 11 ans). L'activité de leur cerveau a été mesurée par électroencéphalographie (EEG) pendant que les enfants effectuaient des problèmes simples d’addition en étant à jeun depuis la veille puis, encore une fois, après avoir mangé ou non un petit-déjeuner. Le nombre de bonnes réponses avait considérablement augmenté chez les enfants ayant mangé, contrairement au nombre de bonnes réponses obtenues chez ceux qui avaient continué de jeûner. Ces résultats donnent à penser que l’activité des réseaux neuronaux traitant les informations numériques et le rendement subséquent sont améliorés chez les enfants ayant pris un petit-déjeuner tandis que ceux qui se passent de déjeuner doivent fournir de plus grands efforts mentaux pour un tel raisonnement mathématique (Pivik et coll., 2012).
Les personnes âgées - Prévenir la détérioration des fonctions cérébrales
Des études transversales ont uniformément montré une détérioration linéaire des fonctions cognitives associées au vieillissement, comme le temps de traitement des données, la mémoire à court terme, la mémoire de travail et la mémoire à long terme. La détérioration des fonctions cognitives liée à l’âge est associée aux changements dans la structure et la fonction cérébrales, et l’activité physique pourrait jouer un rôle déterminant pour ralentir la perte de facultés cognitives avec l’âge (Hillman et coll., 2006). Des méta-analyses récentes portant sur les effets de l’activité physique sur le vieillissement cognitif humain révèlent que les exercices aérobiques ont des d’effets généraux et sélectifs qui améliorent les fonctions cognitives chez les personnes âgées (Colcombe & Kramer, 2003; Etnier et coll., 2006). Ces résultats suggèrent que, même si les facultés cognitives diminuent de façon globale et linéaire avec l’âge, l’activité physique et la capacité aérobique peuvent prémunir contre la perte des facultés cognitives avec l’âge, les plus grands avantages se manifestant pour les processus exigeant un grand contrôle d’exécution. Un nombre grandissant d’études donnent à penser que certains nutriments, comme les flavonoïdes et d’autres polyphénols, peuvent aussi ralentir la détérioration des capacités neuronales et cognitives liée à l’âge (Colcombe et Kramer, 2003; Gómez-Pinilla, 2011).
Structure du cerveau
Il semble que ces résultats se traduisent aussi par des changements structurels au cerveau. Une étude récente montre que le volume de l’hippocampe, une zone du cerveau très importante pour l’apprentissage et la mémoire, est plus important chez les enfants en bonne forme physique chez les enfants du même âge dont la condition physique est moins bonne (Chaddock et coll., 2010a). Une autre étude rapporte que plusieurs autres régions du cerveau ont une structure différente selon la condition physique de l’enfant. Le striatum dorsal qui, croit-on, joue un rôle dans le contrôle et l’inhibition des fonctions cognitives est plus gros chez les enfants en meilleure forme physique (Chaddock et coll., 2010b). Il est possible que les différences de structure observées entre les enfants en bonne et en mauvaise condition physique expliquent, en partie, les différences dans les fonctions cérébrales observées chez les enfants obèses, d'une part, et les enfants ayant un poids santé, d'autre part.
La morphologie du cerveau réagit à des stimuli particuliers pendant toute la vie. Raji et coll. (2010) ont utilisé l’imagerie par résonance magnétique (IRM) pour mesurer l’atrophie de la matière grise et de la matière blanche chez 94 personnes âgées (âge moyen de 77 ans). Les résultats indiquent que l’indice de masse corporelle (IMC), le taux d’insuline à jeun et le diabète de type 2 sont étroitement associés à une atrophie des régions frontales, temporales et sous-corticales du cerveau. Ces données montrent que l'embonpoint ou l’obésité peuvent être associés à de fortes pertes de volume cérébral, et nous permettent de mieux comprendre les causes des changements dus à l'obésité constatés dans la dysfonction cognitive. Étant donné que plusieurs régions du cerveau prédisposées à une perte de volume cérébral sont associées à l’attention, à la mémoire et au contrôle des fonctions cognitives, on peut s’attendre à des déficits sur le plan intellectuel et scolaire en cas d'obésité.
Il semble également que l’entraînement physique chez les aînés peut influer sur la morphologie du cerveau, même à leur âge avancé. Le volume de l’hippocampe et du lobe temporal médial est plus important chez les adultes en meilleure condition physique, et l’activité physique augmente l'apport sanguin de l’hippocampe. Erickson et coll. (2011) ont clairement établi lors d’un essai comparatif à répartition aléatoire mené auprès de 120 personnes âgées (de 55 à 80 ans) qu'un an d'exercices aérobiques augmente la taille de l’hippocampe antérieur, ce qui entraîne une amélioration de la mémoire visuospatiale. L'exercice physique augmente le volume de l’hippocampe de 2 % et peut effectivement reporter d'un an ou deux la perte de volume cérébral due à l’âge. Le volume de l’hippocampe a diminué dans le groupe témoin. Le volume du noyau caudé et du thalamus n'ont pas été touchés par cette mesure. Ces résultats indiquent que les exercices aérobiques peuvent réellement inverser la perte de volume de l’hippocampe à la fin de l’âge adulte, ce qui s’accompagne d’une amélioration de la mémoire (Erickson et coll., 2011).
Mécanismes expliquant les effets positifs de l’exercice sur le cerveau
Des recherches chez l'animal montrent qu’un milieu enrichi, avec des roues d’exercice, a un effet positif sur la croissance des neurones et sur les systèmes neuronaux qui jouent un rôle dans l’apprentissage et la mémoire. La « neuroplasticité » désigne la capacité du cerveau de s’adapter aux changements du milieu environnant, de réagir aux lésions éventuelles et d’acquérir de nouvelles informations en modifiant les connexions et les fonctions neuronales. Les composés favorisant la neuroplasticité s'appellent les « neutrophines» et ils peuvent envoyer des signaux aux neurones afin de survivre, de se différencier ou de croître. Ces facteurs neurotrophiques jouent non seulement un rôle dans la neurobiologie, mais aussi dans le métabolisme énergétique au niveau central et périphérique (Knaepen et coll., 2010). Leur effet sur la plasticité synaptique du système nerveux central (CNS) met à contribution des éléments du métabolisme énergétique cellulaire. L’exercice et un entraînement rigoureux semblent constituer des interventions déterminantes pour déclencher les processus par lesquels les neutrophines induisent le métabolisme énergétique et la plasticité neuronale. Parmi tous les facteurs, le facteur neurotrophique dérivé du cerveau (BDNF) semble le plus sensible à une régulation induite par l’exercice et l’activité physique (Knaepen et coll., 2010). Le BDNF est associé à un vaste répertoire d’activités neurotrophiques et neuroprotectrices dans le CNS et le système périphérique, notamment : la protection et la survie des neurones; l’expression des neurites; la croissance axonale et dendritique, et le remodelage; la différenciation neuronale et la plasticité synaptique comme la synaptogenèse dans l’arborisation des terminaisons axonales et l’efficacité de la transmission synaptique. Des recherches chez l'animal indiquent que le BDNF a aussi une activité neuroendocrinienne ou métabotropique sur le système périphérique. Le BDNF réduit la consommation alimentaire, augmente l’oxydation du glucose, réduit le taux de glycémie et augmente la sensibilité à l’insuline. Chez les animaux, un régime riche en matières grasses réduit la concentration de BDNF dans l’hippocampe, mais l’exercice permet d’inverser cette réduction due à l’alimentation (Molteni et coll., 2004). De plus, une étude chez les souris a montré qu'il y a interaction centrale entre l’hormone leptine dérivée des adipocytes, qui joue un rôle déterminant dans la régulation de l’appétit et du métabolisme énergétique, et l’expression du BDNF dans l’hypothalamus (Komori et coll., 2006). Araya et coll. (2008) ont montré qu’après un régime hypocalorique, les sujets insulinorésistants faisant de l'embonpoint et obèses voient augmenter leur taux sérique de BDNF. Ces résultats confirment que le BDNF est non seulement essentiel pour le système neuronal, mais qu'il est aussi étroitement lié au métabolisme énergétique et à l'homéostase au niveau central et périphérique (Wisse et Schwartz, 2003).
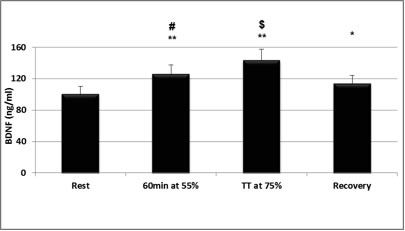
Dans les recherches sur les mécanismes expliquant la plasticité et la santé du cerveau, l’exercice est reconnu comme inducteur d'une série de processus moléculaires et cellulaires qui favorisent la plasticité du cerveau. Le BDNF pourrait jouer un rôle primordial dans ces mécanismes. C’est pourquoi, depuis le début des années 1990, les études ont commencé à analyser les effets de l’activité physique et de l’exercice ou d'un entraînement rigoureux sur la concentration de BDNF, d’abord chez l'animal (Radak et coll., 2006; van Praag, 2009), puis chez l'humain (Gold et coll., 2003; Knaepen et coll., 2010) (Figure 1). On sait peu de choses sur les effets de l'exercice contre résistance sur la mémoire associée à l’hippocampe, même si ce type d’exercice est de plus en plus souvent recommandé pour améliorer la force musculaire et la densité osseuse, et pour prévenir les handicaps liés à l’âge. L’exercice contre résistance augmente le rendement cognitif surtout chez les personnes âgées. Afin d’identifier quels en seraient les mécanismes sous-jacents, Cassillhas et coll. (2012) ont effectué une étude de huit semaines chez l'animal, au cours de laquelle certains sujets ne recevaient aucun entraînement, tandis que d’autres étaient soumis à un programme d’exercices aérobiques et d'autres encore faisaient des exercices contre résistance. Après les huit semaines, lors de leur évaluation dans un labyrinthe aquatique, les facultés cognitives des deux groupes soumis à un exercice physique s'étaient améliorées comparativement à celles des animaux du groupe témoin. Toutefois, différents mécanismes ont été associés aux améliorations cognitives des deux groupes soumis à un exercice physique. Chez les rats soumis aux exercices d’endurance, le BDNF a augmenté de façon significative, tout comme son récepteur, tandis qu'aucune hausse de BDNF n'a été observée chez les animaux entraînés contre résistance. Dans ce dernier groupe, toutefois, le facteur de croissance insulinique de type 1 (IGF-1), une molécule qui stimule la croissance du tissu nerveux et de son récepteur, avait considérablement augmenté. Une analyse plus poussée a révélé que les deux programmes d’entraînement ont engendré une augmentation de l’expression de la synapsine 1 et de la synaptophysine dans le cerveau. Ces deux molécules jouent un rôle important dans la réaction en chaîne nécessaire à la croissance du tissu nerveux et leur augmentation corrobore l’hypothèse selon laquelle les exercices aérobiques et les exercices contre résistance peuvent utiliser des mécanismes moléculaires différents, mais parvenir à des résultats comparables en ce qui concerne l'apprentissage et la mémoire visuospatiale.
INFLUENCE DE L'ALIMENTATION SUR LA FATIGUE DU SYSTÈME NERVEUX CENTRAL
Les interventions en nutrition sont utiles non seulement pour agir sur la fonction cognitive, mais aussi pour prolonger la durée d'un exercice ou retarder la fatigue. La fatigue peut être définie comme une défaillance aiguë des performances sportives engendrant une incapacité à produire une puissance maximale (Meeusen et coll., 2006), ce qui comprend à la fois une augmentation de l'effort perçu pour exercer une force ou une puissance donnée et l'incapacité éventuelle de produire cette force ou cette puissance (Davis et Bailey, 1997). Il se peut que cette fatigue se fasse sentir dans les muscles périphériques ou au niveau du SNC.
Fort probablement, ce sont les neurotransmetteurs cérébraux, et surtout les monoamines centrales, qui stimulent le système nerveux central pour produire les effets de la fatigue lors d’un exercice. Les monoamines comme la sérotonine (5-HT), la dopamine (DA) et la noradrénaline (NA) jouent un rôle primordial dans la transduction de signaux entre les neurones, et il a été établi que les changements de concentration de ces neurotransmetteurs (en particulier la 5-HT et la NA) qui sont induits par l’exercice sont liés à la fatigue centrale. Acworth et coll. (1986) ont d’abord proposé une première hypothèse, élaborée par la suite par Newsholme et coll. (1987), selon laquelle les modifications de la neurotransmission centrale pourraient expliquer la fatigue, et qu'on a appelé « l’hypothèse de la fatigue centrale ».” Cette hypothèse s’appuie sur les écarts de concentration de 5-HT dans le cerveau puisque ce neurotransmetteur joue un rôle dans les changements associés aux périodes de sommeil et d’éveil, les émotions, le sommeil, l’appétit, l’axe hypothalamo-hypophysaire et de nombreuses fonctions physiologiques (Meeusen et coll., 2006). Pendant un exercice, le tryptophane (le précurseur du 5-HT) traverse la barrière hématoencéphalique et pénètre dans le système nerveux central, un processus favorisé par l'utilisation accrue des acides aminés à chaîne ramifiée (AACR) par les muscles et par une concentration plus élevée d’acides gras dans le sang, ce qui augmente le rapport de tryptophane libre aux AACR. Ce processus augmente la quantité de tryptophane qui traverse la barrière hématoencéphalique, ce qui entraîne des taux plus élevés de 5-HT dans le cerveau (Davis et coll., 2000; Meeusen et coll., 2006; Roelands et Meeusen, 2010). Toutes les réactions qui se produisent dans le cerveau peuvent influer sur la façon dont une personne ressent la fatigue et peut donc altérer sa performance. Par conséquent, il est possible de moduler le système nerveux central en modifiant le régime alimentaire ou en y ajoutant des suppléments contenant certains nutriments, dont des acides aminés (AACR, tyrosine), des glucides (CHO) et de la caféine.
Acides aminés à chaîne ramifiée
Il a été suggéré qu'un apport supplémentaire en AACR pourrait permettre de limiter le développement de la fatigue centrale. Si cette théorie semble très attrayante, très peu de preuves directes ou circonstancielles permettent de croire que la performance humaine lors d'un exercice peut être modifiée par des interventions en nutrition, notamment par des suppléments alimentaires comprenant des AACR. À ce jour, aucune preuve convaincante ne vient confirmer que les suppléments d’AACR peuvent retarder la fatigue lors d'un exercice prolongé (Meeusen et Watson., 2007).
Tyrosine
La tyrosine (hydroxyphénylalanine) peut être synthétisée dans l’organisme à partir de la phénylalanine, et plusieurs aliments à forte teneur en protéines, dont les produits du soya, le poulet, la dinde, le poisson, les arachides, les amandes, les avocats, le lait, le fromage, le yogourt et les graines de sésame en contiennent. Une grande consommation de tyrosine augmente son taux par rapport aux autres grands acides aminés neutres comme la leucine, l’isoleucine, la valine et le tryptophane. Une série d’études précliniques chez l'animal ont clairement indiqué que la tyrosine réduit plusieurs des effets indésirables du stress aigu sur le rendement cognitif dans une grande variété de milieux stressants. Même s'il est difficile de démontrer avec certitude que la tyrosine a des effets bénéfiques chez l'humain, en partie en raison de considérations éthiques, la plupart des données probantes indiquent que la tyrosine peut être utile, pour un traitement de courte durée, afin de prévenir une perte de la fonction cognitive liée au stress. D'autant plus si l'exercice se fait par grande chaleur, car le cerveau doit alors produire un supplément de dopamine, ce qui n’est pas le cas dans des conditions tempérées (Roelands et coll., 2008; Watson et coll., 2005). Par conséquent, les besoins du cerveau en tyrosine peuvent s’avérer plus importants en raison des exigences cumulatives de l’exercice et du stress thermique, ce qui pourrait devenir le facteur limitant à la synthèse et à la libération de dopamine. Très récemment, Tumilty et coll. (2011) ont évalué les effets d’une forte supplémentation de tyrosine (150 mg/kg de masse corporelle) sur la capacité à l’exercice par temps chaud. Huit hommes en santé ont pédalé jusqu’à l’épuisement, atteignant 68 ± 5 % du VO2 max. Cette étude révèle, pour la première fois, une association entre un apport nutritionnel en précurseur de la dopamine une heure avant l’exercice et une plus grande capacité à l’exercice par temps chaud. Il semble que la disponibilité de tyrosine peut influer, du moins en partie, sur la tolérance à l'exercice prolongé en présence de stress thermique (Tumilty et coll., 2011). D’autres études s’avèrent nécessaires pour déceler l’influence d’un apport régulier de grandes quantités de tyrosine (5 à 10 g) sur la santé en raison de modifications chroniques dans l’activité du système nerveux sympathique.
Glucides
Plusieurs études ont montré un effet de renforcement par l’exercice de l’apport supplémentaire de glucides sur le métabolisme périphérique; cependant, des études plus récentes suggèrent qu’une alimentation en glucides peut aussi augmenter l’endurance lors d'un exercice en affectant le cerveau, soit par l’effet direct de l’arrivée de glucose au cerveau, soit par les signaux nerveux transmis au cerveau par les récepteurs CHO de la cavité buccale. Carter et coll. ont rapporté une augmentation de 3 % du temps de performance (PLA 61,37 min; CHO 59,57 min) après le rinçage de la bouche avec une solution de maltodextrine avant et après l’exercice. Aucune des solutions n’a été avalée au cours du protocole, ce qui suggère que ce gain de performance peut avoir été favorisé par une communication directe entre le cerveau et les récepteurs de la bouche. Le concept du rince-bouche au CHO s’appuie sur des travaux ayant analysé l’activité cérébrale après l’ingestion d’un bolus de glucose (Liu et coll., 2000), ainsi que l’activation de plusieurs régions du cerveau après rinçage de la bouche avec une solution de CHO (Chambers et coll., 2009). Ces études font ressortir une augmentation marquée de l’activité cérébrale immédiatement après l’arrivée de CHO dans la bouche, avec une deuxième pointe d’activité observée 10 minutes après l’ingestion, laquelle survient sans doute au moment où le substrat entre dans la circulation. D’autres groupes ont également étudié les effets d’un rince-bouche au CHO sur la performance. Pottier et coll. (2010) ont obtenu une amélioration de la performance lors d’une course chronométrée 60 minutes après le rinçage de la bouche avec une solution à base de glucides et d’électrolytes, tandis que les études de Rollo et coll. (2008, 2010, 2011) ont révélé des effets ergogéniques lors de diverses courses chronométrées. Fait à noter, la plupart des études qui ont révélé un effet ont été effectuées quand sujets étaient à jeun. Aucun effet sur la performance n'a été observé lors de courses chronométrées de 45 minutes (Whitham et McKinney, 2007) et de 60 minutes (Beelen et coll., 2009) quand les sujets ayant mangé s'étaient rincé la bouche avec une solution à base de CHO. Les auteurs suggèrent que la perception des glucides dans la bouche pourrait jouer un rôle seulement si les réserves de glycogène dans les muscles et le foie sont réduites. Toutefois, ce résultat n’a pas été reproduit lors d'une étude plus récente analysant le rinçage de la bouche avec une solution contenant de la maltodextrine et au cours de laquelle il y a eu augmentation du temps avant l’épuisement chez des hommes non sportifs (Fares et Kayser, 2011). Ces résultats sont très atypiques et suggèrent un mécanisme d’action intéressant. Une étude plus poussée des récepteurs CHO de la bouche est certainement nécessaire.
APPLICATIONS PRATIQUES ET CONCLUSIONS
L’alimentation fournit les éléments de base à la croissance du cerveau. Le rendement cognitif (raisonnement mathématique et résolution d'additions simples) s’améliore chez les enfants qui ont pris un petit-déjeuner comparativement au rendement obtenu par des enfants à jeun. Un nombre grandissant d’études donnent à penser que certains nutriments, comme les flavonoïdes et d’autres polyphénols, peuvent aussi ralentir la détérioration des capacités neuronales et cognitives liée à l’âge. Chez les personnes âgées, l'exercice physique augmente la taille de l’hippocampe (une région du cerveau qui joue un rôle important dans la mémoire). L’exercice agit favorablement sur les facteurs neurotrophique (comme le BDNF) ce qui améliore l’apprentissage et la mémoire. À l'heure actuelle, aucune preuve convaincante ne vient confirmer que consommer des acides aminés à chaîne ramifiée lors d'un exercice prolongé pourrait retarder la fatigue « centrale ». Il y a aussi un effet ergogénique des glucides pendant un exercice si la bouche est rincée avec une solution à base de CHO.
L’exercice et l'alimentation peuvent tous deux agir de façon efficace et favorable sur le cerveau. Nous commençons seulement à explorer les raisons qui expliquent ce qui se passe réellement dans le cerveau pendant un exercice, mais il est évident que l'activité physique et l'alimentation ont des effets favorables sur le cerveau et la santé. Dans un avenir proche, les interventions en nutrition s'attarderont aussi sur l’activité cérébrale lors d'un exercice.
REFERENCES
Acworth, I., J. Nicholass, B. Morgan, and E.A. Newsholme (1986). Effect of sustained exercise on concentrations of plasma aromatic and branched-chain amino acids and brain amines. Biochem. Biophys. Res. Commun. 137:149-153.
Araya AV, X. Orellana, and J. Espinoza (2008). Evaluation of the effect of caloric restriction on serum BDNF in overweight and obese subjects: preliminary evidences. Endocrine 33:300-304.
Beelen, M., J. Berghuis, B. Bonaparte, S.B. Ballak, A.E. Jeukendrup, and L.J. van Loon (2009). Carbohydrate mouth rinsing in the fed state: lack of enhancement of time-trial performance. Int. J. Sport Nutr. Exerc. Metab. 19:400–409.
Carter, J.M., A.E.Jeukendrup, and D.A. Jones (2004). The effect of carbohydrate mouth rinse on 1-h cycle time trial performance. Med. Sci. Sports Exerc. 36:2107-2111.
Cassilhas, R.C., K.S. Lee, J. Fernandes, M.G.M. Oliveira, S. Tufik, R. Meeusen and M.T. De Mello (2012). Spatial memory is improved by aerobic and resistance exercise through divergent molecular mechanisms. Neuroscience 202:309–317
Chaddock, L., K.I. Erickson, and R.S. Prakash (2010a). A neuroimaging investigation of the association between aerobic fitness, hippocampal volume, and memory performance in preadolescent children. Brain Res. 1358:172–183.
Chaddock, L., K.I. Erickson, and R.S, Prakash (2010b). Basal ganglia volume is associated with aerobic fitness in preadolescent children. Dev. Neurosci. 32:249–256.
Chambers, E.S., M.W. Bridge, and D.A. Jones (2009). Carbohydrate sensing in the human mouth: effects on exercise performance and brain activity. J. Physiol. 587:1779-1794.
Colcombe, S. and A.F. Kramer (2003). Fitness effects on the cognitive function of older adults: a meta-analytic study. Psychol. Sci. 14:125–130.
Davis, C., and S. Cooper (2011). Fitness, fatness, cognition, behavior, and academic achievement among overweight children: Do cross-sectional associations correspond to exercise trial outcomes? Prev. Med. 52:S65–S69.
Davis, J.M., N.L. Alderson, and R.S. Welsh (2000). Serotonin and central nervous system fatigue: nutritional considerations. Am J. Clin. Nutr. 72:573S-578S.
Davis, J.M., and S.P. Bailey (1997). Possible mechanisms of central nervous system fatigue during exercise. Med. Sci. Sports Exerc. 29:45-57.
Erickson K, M. Voss, R. Prakash, C. Basak, A. Szabo, L. Chaddock, J. Kim, S. Heo, H. Alves, S. White, E. Wojcicki, E. Mailey, V. Vieira, S. Martin, B. Pence J. Woods E. McAuley and A. Kramer (2011). Exercise training increases the size of hippocampus and improves memory. PNAS 108:3017–3022.
Rev.
Fares, E.J., and B. Kayser (2011). Carbohydrate mouth rinse effects on exercise capacity in pre- and postprandial states. J. Nutr. Metab. 2011:385926.
Goekint, M., E. Heyman, B. Roelands, R. Njemini, I. Bautmans, T. Mets, and R. Meeusen (2008). No influence of noradrenaline manipulation on acute exercise-induced increase of brain-derived neurotrophic factor. Med. Sci. Sports Exerc. 40:1990-1996.
Gold SM, K. Schulz, S. Hartmann, M. Mladek, U.E.Lang, R. Hellweg, R. Reer, K.M. Braumann, and C. Heesen (2003). Basal serum levels and reactivity of nerve growth factor and brain-derived neurotrophic factor to standardized acute exercise in multiple sclerosis and controls, J Neuroimmunol. 183:99-105.
Gómez-Pinilla, F. (2011). The combined effects of exercise and foods in preventing neurological and cognitive disorders. Prevent. Med. 52:S75–S80.
Hillman, .C, R. Motl, M. Pontifex, D. Posthuma, J. Stubbe, D. Boomsma, and E, de Geus (2006). Physical activity and cognitive function in a cross-section of younger and older community-dwelling individuals. Health Psych. 25:678–687.
Rev.
Knaepen, K., M. Goekint, E. Heyman, and R. Meeusen (2010). Neuroplasticity - exercise-induced response of peripheral brain-derived neurotrophic factor; a systematic review of experimental studies in human subjects. Sports Med. 40:765-801.
Komori. T., Y. Morikawa, K. Nanjo, and E. Senba (2006). Induction of brain-derived neurotrophic factor by leptin in the ventromedial hypothalamus. Neuroscience 139:1107-1115.
Liu, Y., J.H. Gao, H.L. Liu, and P.T. Fox (2000). The temporal response of the brain after eating revealed by functional MRI. Nature 405:1058-1062.
Meeusen, R., and P. Watson (2007). Amino acids and the brain: do they play a role in “central fatigue”? Int. J. Sports Nutr. Exerc. Metab. 17:S37-S46.
Meeusen, R., P. Watson, H. Hasegawa, B. Roelands, and M.F. Piacentini (2006). Central fatigue: the serotonin hypothesis and beyond. Sports Med. 36:881-909.
Molteni, R., A. Wu, and S. Vaynman (2004). Exercise reverses the harmful effects of consumption of a high-fat diet on synaptic and behavioral plasticity associated to the action of brain-derived neurotrophic factor. Neuroscience 123:429-440.
Newsholme, E.A., I. Acworth, and E. Blomstrand (1987). Amino acids, brain neurotransmitters and a function link between muscle and brain that is important in sustained exercise. In: G. Benzi (ed) Advances in Myochemistry. London: John Libbey Eurotext, pp. 127-133.
Parrott, M., and C. Greenwood (2007). Dietary influences on cognitive function with aging from high-fat diets to healthful eating Ann. N.Y. Acad. Sci. 1114:389–397.
Pivik, R., K. Tennal, S. Chapman, and Y. Gu (2012). Eating breakfast enhances the efficiency of neural networks engaged during mental arithmetic in school-aged children. Physiol. Behav.106:548–555.
Pottier, A., J. Bouckaert, W. Gilis, T. Roels, and W. Derave (2010). Mouth rinse but not ingestion of a carbohydrate solution improves 1 h cycle time trial performance. Scan. J. Med. Sci. Sports 20:105-111.
Radak, Z., A. Toldy, Z. Szabo, S. Siamilis, C. Nyakas, G. Silye, J. Jakus, and S. Goto (2006). The effects of training and detraining on memory, neurotrophins and oxidative stress markers in rat brain. Neurochem. Int. 49:387-392.
Raji, C.A., A.J. Ho, N.N. Parikshak, J.T. Becker, O.L. Lopez, L.H. Kuller, X. Hua, A.D. Leow, A.W. Toga, and P.M. Thompson (2010). Brain structure and obesity. Hum. Brain Mapp. 31:353–364.
Roelands, B., H. Hasegawa, P. Watson, M.F. Piacentini, L. Buyse, G. De Schutter, and R. Meeusen (2008). The effects of acute dopamine reuptake inhibition on performance. Med. Sci. Sports Exerc. 40: 879-858.
Roelands, B., and R. Meeusen (2010). Alterations in central fatigue by pharmacological manipulations of neurotransmitters in normal and high ambient temperature. Sports Med. 40:229-246.
Rollo, I., M. Cole, R. Miller, and C. Williams (2010). Influence of mouth rinsing a carbohydrate solution on 1-h running performance. Med. Sci. Sports Exerc. 42:798-804.
Rollo, I., C. Williams, N. Gant, and M. Nute (2008). The influence of carbohydrate mouth rinse on self-selected speeds during a 30-min treadmill run. Int. J. Sport. Nutr. Exerc. Metab. 18:585-600.
Rollo, I., C. Williams, and M. Nevill (2011). Influence of ingesting versus mouth rinsing a carbohydrate solution during a 1-h run. Med. Sci. Sports Exerc. 43:468-475.
Taki, Y., H. Hashizume, Y. Sassa, H. Takeuchi, M. Asano, K. Asano, and R. Kawashima (2010). Breakfast staple types affect brain gray matter volume and cognitive function in healthy children. PLoS ONE 5(12): e15213.
Tumilty, L., G. Davison, M. Beckmann, and R. Thatcher (2011). Oral tyrosine supplementation improves exercise capacity in the heat. Eur. J. Appl. Physiol. 111:2941-2950.
van Praag, H. ( 2009). Exercise and the brain: something to chew on. Trends Neurosci. 32:283-290.
Watson, P., H. Hasegawa, B. Roelands, M.F. Piacentini, R. Looverie, and R. Meeusen (2005). Acute dopamine/noradrenaline reuptake inhibition enhances human exercise performance in warm, but not temperate conditions. J. Physiol. 565:873–883.
Whitham, M., and J. McKinney (2007). Effect of a carbohydrate mouthwash on running time trial performance. J. Sports Sci. 25:1385-1392.
Wisse, B.E., and M. Schwartz (2003). The skinny on neurotrophins. Nat. Neurosci. 6:655-656.